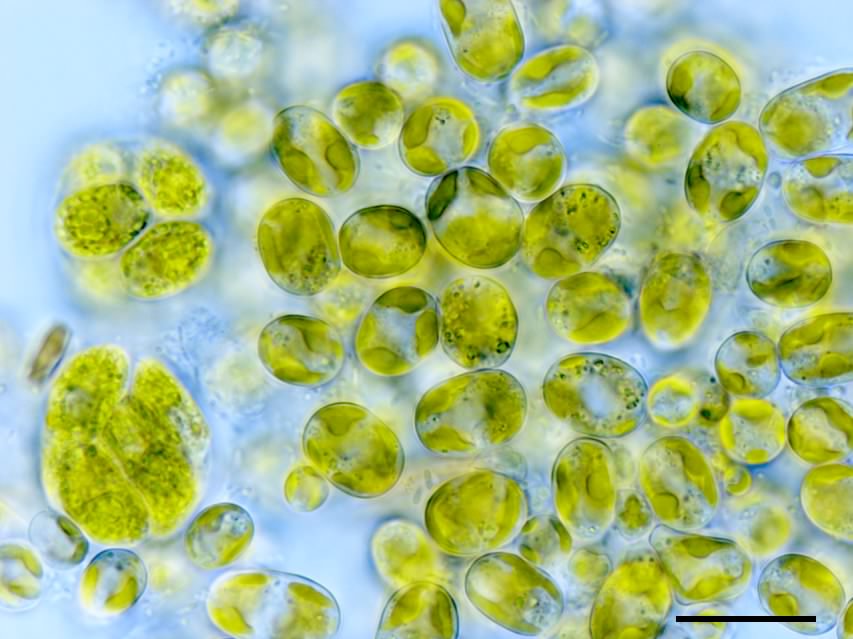
黄緑色藻(おうりょくしょくそう)または黄緑藻(おうりょくそう)(英: yellow-green algae, xanthophytes)とは、不等毛藻(オクロ植物門)の1綱である黄緑色藻綱(黄緑藻綱、学名: Xanthophyceae)のこと、またはこれに属する生物のことである。およそ90属600種ほどが記載されている。多くは単細胞性や糸状性の微細藻であるが、フウセンモ(Botrydium; 図1)やフシナシミドロ(Vaucheria)は肉眼視できる大きさの多核体をもつ。不等毛藻としては例外的に葉緑体は緑色であるが、これは不等毛藻に一般的なカロテノイドであるフコキサンチンを欠くためである。そのため緑藻に似るが、デンプンをもたないことや、鞭毛細胞が不等鞭毛をもつことなどで区別できる。主に淡水域に見られるが、陸上域や海に生育するものもいる。一部の種は真正眼点藻綱に移されたが、他にも移されるべき種が多く残されていると考えられている。
特徴
体制
黄緑色藻の体制(体のつくり)は多様であり、単細胞性のものから群体性、糸状性、多核嚢状性のものが知られている。単細胞性のものは基本的に細胞壁に囲まれた不動性であり、球形のもの(ボトリディオプシス属 Botrydiopsis)や円筒状のもの(オフィオキチウム属 Ophiocytium; 下図2a)、紡錘形で基物に付着しているもの(カラキオプシス属 Characiopsis; 下図2b)などがある。群体性のものの中には、寒天質に包まれたパルメラ状群体のもの(下図2c)から、細胞が密着したサルシナ状群体を形成するもの(下図2d)がある。またミスココックス属(Mischococcus)は、分岐する寒天質の柄のそれぞれの先端に球形の不動細胞が存在する。糸状体のものは、トリボネマ属(Tribonema)のように分枝しない単列糸状のもの(下図2e)や、ヘテロコックス属(Heterococcus)のように分枝するもの(下図2f, g)がある。フウセンモ属(Botrydium)は直径数ミリメートルに達する多核嚢状部と、そこから地中に伸びる分枝した仮根部からなり、隔壁がない(下図2h, 3a)。フシナシミドロ属(Vaucheria)は隔壁がなくまばらに分枝する糸状体からなり(下図2i)、しばしば肉眼視できるマットを形成する。
黄緑色藻の中には、単細胞鞭毛性やアメーバ性の種も報告されている(下図5a, b)。しかし、このような種の中で詳細な研究から黄緑色藻に属することが確認された例はない。
細胞構造
黄緑色藻の細胞は、単核性(核は1個)のものから多核性(核は多数)のものまで存在する。核分裂の過程が詳細に調査された例は少ないが、中心体が両極に位置し、極も含めて完全な閉鎖型(核分裂中に核膜が崩壊せずに維持される)である核分裂が報告されている。
ふつう複数のゴルジ体が核に面して存在するが、フシナシミドロ属ではゴルジ体が小胞体 (ER) とミトコンドリアに密接している。
葉緑体は緑色、ふつう盤状であり、細胞外縁に沿って1個から複数存在する。葉緑体の最外膜は、しばしば核膜と連続している。チラコイドは、ふつう3枚ずつ重なってチラコイドラメラを形成している。ふつう周縁ラメラ(ガードルラメラ)をもち、色素体DNAはその内縁に沿ってリング状に存在するが、周縁ラメラを欠き色素体DNAが分散しているもの(Bumilleria, Bumilleriopsis など)もいる。
葉緑体は多くの緑藻に見られるような明瞭なピレノイドを欠くが、詳細な観察からは多くが半埋没型(葉緑体表層に位置し細胞内側へやや突出している)ピレノイドをもつことが示されている。このようなピレノイドの基質には、やや離れて平行に並んだチラコイドラメラが横断している。他の不等毛藻で報告されているような、ピレノイド外面を覆うキャップ状小胞は報告されていない。ただし、脂質顆粒(plastoglobuli)がピレノイド表層に集まっていることがある。また、Tribonema viride や Vaucheria woroniniana では、基質中にチラコイドが貫入していない突出型のピレノイド様構造が報告されているが、この構造が実際にピレノイドに相当するのか否かは明らかではない。
クロロフィルとしてはa、c1、c2をもつが、クロロフィルc量は極めて少なく、a:c比は 55:1から116:1ほどであることが報告されている。古くにクロロフィルeが報告されたことがあるが、その後見つかっていない。カロテノイドとしては、ジアジノキサンチン、ジアトキサンチン、ヘテロキサンチン、ボウケリアキサンチンエステル、β-カロテンなどをもつ。一部の種では、ネオキサンチンやクリプトキサンチンモノエポキシドなどが少量報告されている。他の不等毛藻に一般的なフコキサンチンを欠くため、葉緑体は緑色から黄緑色を呈する。高濃度の塩酸で処理することによって、一部の種(全てではない)では、青緑色を呈することが知られているが、これはキサントフィルの変化によるものであると考えられている。ただし、Pleurochloridella などはフコキサンチンをもつことが報告されており、ファエオタムニオン藻綱に分類されたが、系統的には黄緑色藻の基部に位置しており、黄緑色藻綱に分類されることもある。
貯蔵多糖は他の不等毛藻と同様にβ-1,3/1,6グルカンであるクリソラミナリンであり、小胞中に貯蔵される。葉緑体の色調がよく似た緑藻とは異なり、デンプン(α-1,4グルカン)を貯蔵しないため、ヨウ化カリウム溶液で染色されない。また、細胞質中に脂質を比較的多く貯蔵する。
細胞壁
黄緑色藻の細胞は、ふつう細胞壁で囲まれている。一般的な細胞壁に見られるように細胞壁がひとつながりである場合もあるが、一部の黄緑色藻では、半球形から円筒形で次第に薄くなる縁辺で重なり合った2つのパーツからなる細胞壁をもつ。ただし、通常の状態ではこの特徴が明瞭ではなく、染色(メチレンブルーやコンゴーレッドなど)やアルカリ溶液(水酸化カリウムや水酸化ナトリウム)による細胞壁の変性、遊走子放出による母細胞の裂開などによって初めて2つのパーツからなることが認識されることもある。この2つのパーツは同形同大である場合から、極端に大きさが異なる場合まであり、またトリボネマ属(Tribonema)など糸状性の種では隣接する細胞のパーツ同士が合着して縦断面でH形の構造となっている(上図2e)。
多くの黄緑色藻(全てではない)の細胞壁は、酸による加水分解処理なしでもシッフ試薬によって強い陽性反応を示すことが知られているが、その理由は明らかではない。細胞壁の組成が研究された例は少ないが、細胞壁にセルロースが含まれることが報告されている。一方で上記のような2つのパーツからなる細胞壁は、主成分が酸性ペクチンであるとされることがある。また、珪酸質を含むともされる。
鞭毛細胞
黄緑色藻の多くは、生活環の一時期に遊走子などの形で(下記参照)鞭毛をもつ細胞を形成する。鞭毛細胞は、細胞亜頂端から生じて前後に伸びる2本の不等鞭毛をもつ(上図2b, c, d, f)。一般的に前鞭毛の方が長いが(細胞長と同程度)、フシナシミドの精子では後鞭毛の方が長い。他のストラメノパイルと同様、前鞭毛には管状小毛が付随している。管状小毛の先端毛は2本。ふつう後鞭毛の基部には鞭毛膨潤部があり、1層の色素顆粒からなる葉緑体内の眼点に相対している。鞭毛と基底小体の移行部には、1重または2重のらせん構造 (transitional helix) が存在する。鞭毛細胞には、ふつう2個の葉緑体が細胞の背腹に配置しているが、腹側のみに1個有するものもある。フシナシミドロの精子は葉緑体を欠き、また眼点も存在せず、走光性を示さない。またフシナシミドロの精子は、細胞前端にプロボスシス (プロボーシス、proboscis) とよばれる舌状の突出構造があり、微小管性鞭毛根の一部 (root 3) が発達したもので支えられている。核は腹側葉緑体に接して存在し、その前方に1個のゴルジ体、さらに前方に収縮胞(浸透圧調節のための構造)がある。また、鞭毛細胞の細胞質にはふつう内容物を含むさまざまな小胞が存在し、着生や細胞壁形成に関わっていると考えられている。
フシナシミドロ属の遊走子(集合遊走子)は大型で多数の鞭毛、核、葉緑体を含む特異な形態をしており、いずれも管状小毛を欠く2本の亜等長の鞭毛が組になり、細胞全面から生じている(下図3b左)。眼点をもたず、走光性を示さない。鞭毛組の基部には、それぞれ核が存在する(下図3b左)。
ゲノム
黄緑色藻のミトコンドリアでは、遺伝コードが他とはやや異なり、AUAがメチオニンをコードしている(標準ではイソロイシン)。
生殖
一般的に遊走子(zoospore; 鞭毛をもつ胞子)や自生胞子(autospore; 母細胞と同じ形態をした胞子)によって無性生殖を行う。また、不動胞子(aplanospore; 遊走子に似るが鞭毛をもたない胞子)を形成する例も知られている。糸状性の種は細胞分裂によって成長し、分断化によっても無性生殖を行う。フシナシミドロ属は、糸状体の先端部の原形質が切り離されて多核多鞭毛の遊走子(集合遊走子、集合性遊走子 synzoospore)を形成し、これによって無性生殖を行う(下図3b左)。
鞭毛性やアメーバ性の種は、二分裂によって無性生殖を行う(ただし下記のように、これらの種の黄緑色藻への所属は疑問視されている)。また、このような種の中には、内生的に細胞壁を形成してシストとなるものも報告されている。
有性生殖が知られている例はわずかである。フウセンモ属では、2本鞭毛性の同形または異形配偶子の接合による接合子形成が報告されている(上図3a)。フシナシミドロ属は特別な有性生殖器官を形成し、生卵器で形成された卵と造精器で形成された2本鞭毛性の精子の間での卵生殖を行う(上図3b右)。フシナシミドロ属の接合子(卵胞子 oospore)は厚い細胞壁をもち耐久細胞として機能し、発芽して栄養体を形成する際に減数分裂を行うとされていたが、配偶子形成時に減数分裂を行うことが示唆されている。
フシナシミドロの卵胞子(上記)を除き、黄緑色藻では耐久細胞の報告は少ないが、一部の種で細胞壁が厚化してアキネート (akinete) を形成することが知られている。また一部の種では、2つのパーツからなる細胞壁で囲まれた特殊な胞子を形成することが報告されている。
生態
黄緑色藻の多くは淡水止水域に生育し、特に高層湿原など低pHの環境に多い。ふつう個体数は少なくまれな存在であり、目に触れることは少ないが、トリボネマ属(Tribonema)は大増殖して水面に藻塊を形成することがある。土壌に生育するものも少なくないとされ、南極などからも見つかっている。また、雪上からも報告されている。フウセンモ属(Botrydium)は、湿土上に多く見られることがある。フシナシミドロ属(Vaucheria)は、淡水、汽水、海水域の泥上や陸域の土壌上に生育しており、場所によってはマットを形成している。
嚢舌類ウミウシの一種であるエリシア・クロロティカ(Elysia chlorotica; 下図4a)は、卵から孵化すると塩湿地に生育する黄緑色藻である Vaucheria litorea を食べ、その葉緑体を保持して自らの細胞器官(盗葉緑体)とし、その後は一生光合成を行って生きる。
Heterococcus は、地衣類のアナイボゴケ属(Verrucaria; 子嚢菌門ユーロチウム菌綱)の共生藻となることがある。
Trypanochloris clausiliae は、陸生のキセルガイの殻上のみから報告されている(上図4b)。
系統と分類
19世紀には、葉緑体の色調の類似性などから、当時知られていた黄緑色藻は緑藻に分類されていた。しかし19世紀の終わり頃から、明瞭なピレノイドを欠く盤状で黄緑色の葉緑体をもち、貯蔵物質としてデンプンを欠き脂質を有し、2個の側性葉緑体と1本の鞭毛(実際には前後に伸びる2本の不等鞭毛であるが、当時は誤ってこのように考えられていた)をもつ鞭毛細胞を形成する藻群として認識されるようになった。当初は Confervales として緑藻の一群に分離されていたが、やがて独立の分類群として扱われるようになり、不等毛綱(Heterokontae)や黄緑色藻綱(黄緑藻綱、Xanthophyceae)の名が提唱された。やがて20世紀前半には、アドルフ・パッシャー (Adolf Pascher) によって多数の黄緑色藻(2017年現在知られている属のおよそ2/3)が記載された。
およそ90属600種ほどが記載されている。ただし上記のようにほとんどの黄緑色藻は比較的まれな存在であり、原記載以降、報告がないものも多い。特に鞭毛性のもの(クロラモエバ目; 下図5a)やアメーバ性のもの(リゾクロリス目; 下図5b)の中で、確実に黄緑色藻綱に属することが示された例はない。アメーバ性の Chlorarachnion(下図5c)や Chlamydomyxa は古くは黄緑色藻綱に分類されていたが、その後の研究でこれらは黄緑色藻ではなく、前者はケルコゾア門クロララクニオン藻綱、後者はオクロ植物門シンクロマ藻綱に属することが明らかとなっている。
また、黄緑色藻綱に分類されていた藻類の一部(Vischeria, Chlorobotrys, Goniochloris (上図5d), Pseudostaurastrum など)は、細胞の微細構造や鞭毛細胞の形態、光合成色素組成などの点で典型的な黄緑色藻とは異なることが明らかとなり、真正眼点藻綱(Eustigmatophyceae)として分けられた。真正眼点藻綱は、黄緑色藻綱と同じくオクロ植物門(不等毛植物門)に属するが、その中では系統的に近縁ではない。また、黄緑色藻の中には、真正眼点藻綱に移すべき藻類がまだ残されていると考えられている。
黄金色藻(一部は黄緑色藻)に分類されていたいくつかの藻類が、系統的にこれらとは異なることが示され、新たに設立されたファエオタムニオン藻綱に分類された。設立当時からファエオタムニオン藻綱には2つの系統群(ファエオタムニオン目、プレウロクロリデラ目)が存在することが知られていたが、その後の研究から後者は黄緑色藻の姉妹群であることが示されている。そのため、このプレウロクロリデラ目(Phaeobotrys, Pleurochloridella を含む)を黄緑色藻綱に分類していることもある。
上記の鞭毛性、アメーバ性のものを含め、黄緑色藻はおおよそその体制に基づいて分類されている(下表1)。ただし、分子系統学的研究からは、この意味での分類群の多くが単系統ではないことが示されており(下図6)、この分類体系は系統関係を反映したものではない。いくつか(Botrydiopsis, Chlorellidium)は属レベルでも単系統群ではないことも示されている。しかし、上記のように黄緑色藻の多くはまれにしか見つからないため、詳細に研究された種は限られており、系統関係を反映した分類体系の再構築には至っていない(2023年現在)。フシナシミドロ目とそれ以外(トリボネマ目の名を用いている)の2目に分けた体系もあるが、この体系も明らかに系統関係を反映していない。
脚注
注釈
出典
関連項目
- 不等毛藻(オクロ植物門): 珪藻、ボリド藻、ディクティオカ藻、ペラゴ藻、真正眼点藻、ピングイオ藻、シンクロマ藻、黄金色藻、ラフィド藻、ファエオタムニオン藻、ファエオサッキオン藻、褐藻
外部リンク
- “Xanthophyceae”. Algaebase. 2024年3月30日閲覧。(英語)
- “黄緑藻植物”. 原生生物情報サーバ. 2024年4月5日閲覧。
- “黄緑色藻”. ねこのしっぽ -小さな生物の観察記録-. 2024年4月5日閲覧。(真正眼点藻を含む)