カブトガニ類(英語: xiphosuran, horseshoe crab, 学名: Xiphosura)または剣尾類(けんびるい)は、鋏角類に分類される節足動物の分類群(目、亜綱、もしくは綱)の1つ。ドーム状の甲羅と剣のような尾をもつ。構成種は一般にカブトガニ(兜蟹、鱟)と総称されるが、これは本群の1種 Tachypleus tridentatus を指す和名でもある。鋏角類であり、すなわち名前に反してカニなどの甲殻類ではない。
現生のカブトガニ類は4種のみ知られるが、絶滅した化石種まで範囲を広げると80種以上をも含んだ大きなグループである。カブトガニ類はオルドビス紀で既に出現し、正式に記載されたものはおよそ4億4,500万年前(ヒルナント期)、未命名の化石まで範囲を広げるとおよそ4億8,000万年前(トレマドッグ期)まで遡れる。また、現生のカブトガニ類は一部の化石種によく似た姿をもつため、「生きている化石」の代表的な生物として知られている。
カブトガニ類の分類の内部構成については、従来通りハラフシカブトガニ類(Synziphosurina)を含んだ「広義のカブトガニ類(Xiphosura sensu lato)」と、2013年で再定義され、それを除いた「狭義のカブトガニ類(Xiphosura sensu stricto)」がある。本項目では主に前者について扱う。
なお、カブトガニ類と大まかに似た姿で同じ「生きている化石」として知られ、しばしばカブトガニ類と混同されるカブトエビは甲殻類であり、鋏角類であるカブトガニ類とは別物である。
名称
学名「Xiphosura」はギリシャ語の「Xiphos」(剣)と「uros」(尾)の合成語で、剣のような尾節に因んで名づけられた。
カブトガニ類は鋏角類であるため、カニどころか甲殻類ですらない。しかし、各地域のカブトガニ類への呼称に「カニ」(蟹、crab)を含むことが多く、以下の例が挙げられる。
- 和名は「カブトガニ」と言い、これは日本に分布する唯一のカブトガニ類 Tachypleus tridentatus の標準和名でもある。漢字転写は「鱟」・「兜蟹」などがある。また、別名として「剣尾類」(けんびるい)がある。
- 英語では蹄鉄(horseshoe)の形に似た背甲から「horseshoe crab」の他に、「king crab」や、本群の学名 Xiphosura に因んだ「xiphosuran」という学術的な総称もある。
- 中国語では「鱟」(簡体字: 鲎、ピンイン: Hòu, ホウ)・「馬蹄蟹」(英名「horseshoe crab」と同義)・王蟹(英名「king crab」と同義)の他に、雌雄が前後をくっつける繁殖行動から「夫妻魚」や「鴛鴦魚」とも呼ばれる。
形態
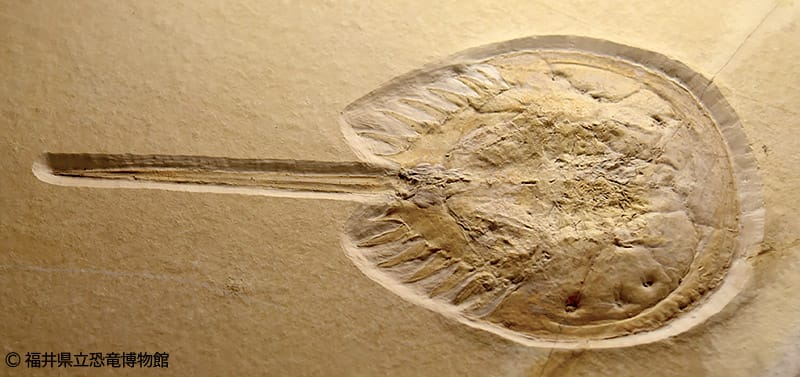
カブトガニ類の身体は、前体と後体の2部によって構成され、末端は剣に似た尾節を持つ。全ての付属肢(関節肢)は腹側にあり、幅広い甲羅状の体に覆われている。現存種はどれも数十cmに及ぶ大型の節足動物であるが、絶滅した化石種まで範囲を広げると数cm程度の小型種の方が多い。また、現存種の外見はお互いによく似ているが、化石種、特に中生代のものは比較的に多様化していた。
前体
前体(prosoma)は先節と第1-6体節の融合でできた合体節である。背面の外骨格、いわゆる背甲(carapace, prosomal dorsal shield)は幅広い半円形から三日月形に発達し、全ての腹面構造を覆いかぶさったドーム状の甲羅となる。背面の左右と中央は、原則として各1対の複眼(側眼 lateral eye)と単眼(中眼 median ocelli)がある。ただし現存種の場合、複眼の直後・腹側の口の前という2箇所からそれぞれ1対の特殊な眼(larval eye)の存在が確認でき、そのうち腹側のものは腹眼(腹側眼 ventral eye)とも呼ばれる。アメリカカブトガニの場合は3つ(通常の1対とそのやや後ろ中央に1個)の中眼をもつ。ハラフシカブトガニ類の場合、ほとんどの種類は眼が退化消失したと考えられる。
カブトガニ類の背甲は、眼以外にも様々な特徴をもつ。複眼の上方には眼部隆起線(opthalmic ridge)が走り、後縁中央から心葉(cardiac lobe, または心域 cardiac region)という盛り上がった領域が前へ伸びて、両後側から突き出した部分は後角(posterior angle)、頬棘(genal spine)、または側棘(lateral spine)と言う。腹面は縁に沿って折り返した外骨格構造(doublure)があり、これと背甲の境目、すなわち背甲の縁辺部は脱皮の割れ目でもある。これらの特徴は分類群によって形や発達具合が異なり、カブトガニ類の系統関係を示すのに重要視される形質である。
前体第1-6体節に由来する6対の付属肢は全て腹面の中央付近で接しており、口はその間に後方に向かって開いている。口と上唇(labrum)の前にある第1対の付属肢は、摂食用の短い鋏型の鋏角(chelicera)である。次の5対は脚で6節以上に分かれ、先端2節が鋏をなしているものが多い。そのうち最初の1対は触肢(pedipalp)であるが、クモガタ類ほどには特化せず、他の脚との区別はほぼない。ただし、少なくともカブトガニ科の雄の場合、この付属肢1対のみ、もしくは次の脚と共に2対が、雌を包接するため先端が太い鉤爪状の構造(把持器 clasper)に特化した。現生種の場合、最終1対の脚は最も発達で、最初の肢節(基節 coxa)は「櫂状器」(flabellum)という葉状の(外肢 exopod もしくは外葉 exite とされる)分岐を上方に有し、最終肢節の付け根周りに「へら状器」(spatulate organ)という4枚の平たい構造体を持つことが確認できる。また、それぞれの脚の基節は、口を囲んだ顎基(gnathobase)という摂食用の鋸歯状の突起(内葉、内突起)がある。脚の間と口の直後には「endostoma」という、腹板(sternite)由来と思われる目立たない外骨格が1枚ある。
クモガタ類と同様、カブトガニ類の前体も便宜的に「頭胸部」(cephalothorax)と呼ばれることがあるが、合体節的には頭部と胸部/胴部の融合ででき頭胸部ではなく、頭部のみに該当する頭部融合節である。ただしカブトガニ亜目の場合、背甲は前体だけでなく、(他の節足動物の胴部に該当する)後体の一部まで含まれるため(詳細は後述)、背甲部分なら頭胸部とも言える。
後体
後体(opisthosoma)は便宜的に「腹部」(abdomen)ともいうが、合体節的には胴部に該当する。種類によって9から11節(第7から15/17体節)を含め、多くの場合は前体より横幅が狭い。カブトガニ亜目の場合、後体前方の体節は複雑な遊離と融合が進み、第1節(第7体節)と第2節(第8体節)の中央まで前体と融合して背甲に覆われ、遊離した後体第2節の縁辺部は「free lobe」という出っ張りとなって後体第3節(第9体節)以降の全ての体節と融合し、「thoracetron」という1枚の甲羅をなしている。ただし基盤的な化石群、いわゆるハラフシカブトガニ類の後体は全ての体節が可動な背板(tergite)に分かれている。背板は三葉状、すなわち中央1対の縦溝で正中線の軸部(axial region)と左右に出張った肋部(tergopleura)に分けられる。ただし多くのカブトガニ亜目の種類はそれが不明瞭で、主に対になる窪み(内突起 apodeme)や棘(epimera)によって現れる。カブトガニ亜目の中で、thoracetron の両脇に後体第3-8節(第9-14体節)由来の縁棘(marginal spine)と呼ばれる6対の可動棘(movable spine)を epimera の間に持つ例が多い。
カブトガニ類の後体第1節(第7体節)は、多くが外見上から認められないほど退化的で、現存種の場合、この体節は前述の通り前体の背甲と融合し、腹面は唇様肢(chilarium)という、最終の脚の間に配置される短い付属肢を持つ。しかし、ほとんどの化石種の第7体節/付属肢の詳細は不明で、一部のハラフシカブトガニ類においては独立した退化的な背板(microtergite)によって現れる。中でもウェインベルギナは、この部分が前体のとほぼ同形な脚を持っていたと考えられる。これらの性質により、この体節は機能的にむしろ前体的で、前体の一部と扱うべきことも検討される。
次の後体第2-7節(第8-13体節)は、6対の蓋板(がいばん、operculum)という平たい付属肢を持ち、基本的には幅広い外肢と指のような短い内肢(endpod)からなる二叉型で、両付属肢の基部中央には「sternal lobe」という、腹板由来と思われる小さく尖った構造体がある。最初の1対は左右融合した生殖口蓋(genital operculum)と言い、卵や精子を排出する1対の生殖孔(gonopore)がその後ろ側の基部にある。次の5対は鰓脚(branchial appendage または gill operculum)で、本のページを彷彿とさせる呼吸用の書鰓(しょさい、book gill)は、それぞれの外肢の後ろ側からぶら下がるように生えている。これらの蓋板は普段では後ろ向きに畳まれるため、鰓脚は生殖口蓋によって覆われて守らていれる。
後体の末端に当たる肛門の直後は、1本の棘状の尾節(telson)が伸び、これは「尾剣 」や「cadual spine」とも呼ばれる。断面は三角形(三角柱状)もしくは円形(円柱状)で、付け根の関節を介して上下と左右に動ける。カブトガニ亜目の尾節は往々にして全長の半分以上を占めるが、ハラフシカブトガニ類の尾節はそれに対しやや短い。
内部構造
液体状の物質しか摂食できない他の多くの現生鋏角類とは異なり、カブトガニ類は固形状の食物を摂ることに適した特殊な消化器を持つ。
前体腹側の口は後方に向かって開き、クチクラに覆われる食道はそこから前方まで伸びる。前体の前縁付近には、Uターンのように、食道から上向きに大きく曲がり返した前胃がある。前胃は素嚢と筋肉の発達した砂嚢からなり、食物を細かく砕き、殻や骨などの食べられない物質はここから口へ噴き返す。砂嚢の直後は細くなり、幽門弁に続く。幽門弁から肛門付近の直腸までの消化管は長い中腸でつながっており、その左右は体の大部分を占めた盲腸(中腸腺)がある。消化酵素はここで分泌され、食物を分解し、栄養を吸収する。
循環系は他の鋏角類よりも高度に発達しており、背側の縦長い心臓や体の縁まで枝分かれ数多くの動脈を含んだ血管系と、その左右と書鰓に繋いだ間隙循環系(lacunar circulatory system)でできている。体内の血リンパは無色透明であるが、ヘモシアニンを含むため、流血などで酸素と結びつくと銅イオン由来の青色になる。
他の節足動物と同様、カブトガニ類ははしご形神経系を持ち、脳は前大脳・中大脳・後大脳と呼ばれる3つの脳神経節からなるが、節足動物として原始的と思われる形質を持つ。また、前体の神経節はクモガタ類のように集約する(synganglion をなす)が、後体の神経節は明瞭に分節している。
排出器官として1対の基節腺(coxal gland)があり、開口部は後脚の基節に開く。これもクモガタ類に似ている(触肢・第1脚・第3脚のいずれかの基節に開く)。
生態
現存種のカブトガニ類は干潟や海底に生息する底生性の海棲動物である。肉食偏りの雑食性で、確保した餌は脚の顎基の連動によって握りつぶされ、唇様肢で漏れることを防ぎつつ、鋏角でそれを口へと運ぶ。二枚貝を主食とし、巻貝・多毛類・甲殻類・腐肉・海藻など様々な生物をも餌とする。後脚のへら状器と幅広い背甲を利して、干潟や砂浜の上でも泥に囚われずに前進できており、干潟上の移動や、砂などの堆積物の中に潜むことに適している。後体の蓋板を前後に羽ばたいて、裏返しの姿勢で泳くこともできる。成体の性的二形は明瞭で、雌に比べて雄はやや小柄であり、包接中に雌の後体を噛合せるように、背甲の前縁下側もやや上方にくぼんでいる。繁殖期の頃、干潟や砂浜に集まり、雄は特化した鉤爪状の歩脚を用いて雌の後体を包接する。雌は砂浜上で数万個の卵を産み、雄は排精して卵の体外受精を完成させる。幼生は十数回の脱皮をし、数年を経て成体になる。節足動物にしては長寿で、例えばアメリカカブトガニの平均寿命は20年から40年だと推測されている。長寿である上に、成体になると脱皮が止まるため、大きな体にはフジツボや巻貝などの付着生物が付くことが多い。
なお、絶滅した化石種は現生種以上に多様で、淡水域に生息・遊泳や堆積物に潜むことに不向き・全身を丸めるなど、現存種とは異なった生態や行動を持つと考えられる種類までいくつか知られている。特に石炭紀で繁栄したヒメカブトガニ類(Bellinuridae)は、その全ての種類が淡水生であったと考えられる。
現存種
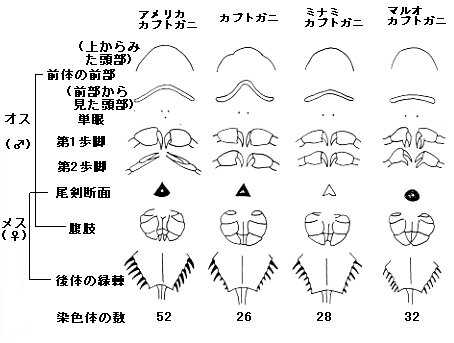
現生のカブトガニ類は、カブトガニ(Tachypleus tridentatus)、ミナミカブトガニ(Tachypleus gigas)、マルオカブトガニ(Carcinoscorpius rotundicauda)、アメリカカブトガニ(Limulus polyphemus)の4種のみが知られる。各種類の詳細はそれぞれの該当項目を参照のこと。
- カブトガニ Tachypleus tridentatus
- 日本、台湾、中国沿岸など東アジアと東南アジアの海域に生息する。体長は雄45cmから70cm、雌55cmから85cmほどまで成長し、知られる中では最大のカブトガニ類である。
- ミナミカブトガニ Tachypleus gigas
- 南アジアと東南アジアの海域に生息する。体長は雄25cmから40cm、雌25cmから50cm程度である。
- マルオカブトガニ Carcinoscorpius rotundicauda
- 南アジアと東南アジアの海域に生息する。大体長は雄35cm、雌40cm、現存種の中では最小のカブトガニ類である。時期によって毒を持つことが知られている。
- アメリカカブトガニ Limulus polyphemus
- アラスカ州西部と、メキシコ湾など大西洋北西沿岸の海域に生息する。現生カブトガニ類の中で研究が最も進んでいる種である。
いずれも生態と外見はよく似て、生息地と体長の相違点以外では以下の特徴から区別できる。
分類
分類史
19世紀には大まかな類似に基づいて、節足動物の中でカブトガニ類は特に三葉虫に近縁と考えられた。当時はウミサソリ類と共に、いずれも甲殻類扱いされた。しかし20世紀以降の再検討により、カブトガニ類とウミサソリ類は甲殻類らしからぬ、むしろサソリなどのクモガタ類に共通な基本体制を持つことが判明した。それ以降、三葉虫や甲殻類と区別されたカブトガニ類とウミサソリ類は、ウミグモ類・クモガタ類などと共に、まとめて節足動物の大グループの1つである鋏角類(Chelicerata)に含まれるようになった。
系統関係
カブトガニ類は、複眼・顎基・二叉型付属肢・発達した後体付属肢と書鰓など、残りの現生真鋏角類(ウミグモ類以外の鋏角類)であるクモガタ類(クモ・サソリ・ダニなど)には見当たらない、真鋏角類の祖先形質と思われる特徴が数多く整っているため、真鋏角類の初期系統発生およびその共通祖先への考査において豊富な情報源として重要視される分類群である。通常、派生的と思われるクモガタ類に対して、カブトガニ類はそれより早期に分岐した基盤的な真鋏角類と考えられる。カブトガニ類とクモガタ類に至る系統は、少なくとも古生代オルドビス紀の頃に分岐していたと考えられる。
なお、一部の分子系統解析では別の系統関係を示唆し、カブトガニ類は非単系統群のクモガタ類の間から分岐し、その中でクツコムシ類もしくは蛛肺類(書肺のあるクモガタ類、クモ・サソリ・ウデムシ・サソリモドキなどを含む)に近縁という解析結果を出していた。これを踏まえて、カブトガニ類は陸生のクモガタ類に起源で二次的に水生化した、もしくはカブトガニ類が水生のままでクモガタ類が複数回で上陸したという2つの説も提唱された。もしカブトガニ類は蛛肺類に近縁であれば、カブトガニ類と蛛肺類の全ゲノム重複は両者の共有派生形質かもしれないが、もしそうでなければ、これは収斂進化の結果と見なされる。
しかし、このような見解(特に二次的水生化説)に対する反発が多く、中には通説(カブトガニ類は単系統群のクモガタ類よりも早期に分岐)を支持する分子系統解析と分子時計的解析や、カブトガニ類の二次的水生化説を否定する古生物学的証拠が挙げられた。特に後者に関して、水生性に適した特徴(顎基・書鰓など)を含めてカブトガニ類の多くの性質は、クモガタ類以外の化石鋏角類(ウミサソリ類など、後述参照)を通じて広く見られ、祖先形質であることが強く示唆される。例えクモガタ類は本当にカブトガニ類に対して非単系統だとしても、最大節約法的にクモガタ類が複数回で上陸した説の方が現実的であり、カブトガニ類の水生性質はおそらく祖先形質で、とても二次的には考えられない。
化石鋏角類との関係性
カブトガニ類に類するほどの現生鋏角類は存在しないが、絶滅した化石鋏角類の分類群まで範囲を広げると、ウミサソリ類、カスマタスピス類、および広義のカブトガニ類に含まれるハラフシカブトガニ類など多く知られ、カブトガニ類との関係性は多く議論を繰り広げられた。カブトガニ類とこれらの群は、水生性・発達した背甲・書鰓のある後体付属肢・複眼・顎基のある前体付属肢など多くの共有形質を有し、古典的にはクモガタ類と対になる節口類(Merostomata, 腿口類とも)としてまとめられていた。しかしこの分類体系の単系統性は20世紀後期から徐々に疑問視され、節口類全体がモガタ類に対して側系統群で、すなわち前述の共通点は真鋏角類の祖先形質という見解が主流となっている。この見解を踏まえて、節口類は単に便宜的な総称となり(もしくはカブトガニ類以外の群を節口類から除外し、節口類 Merostomata をカブトガニ類 Xiphosura のシノニムとする)、カブトガニ類を節口綱に内包される目から独自の綱へ昇格させる分類体系も採用されつつある。節口類の単系統性を支持する解析結果もわずかにあるが、この場合の節口類は基盤的な鋏角類、もしくはクモガタ類が節口類に対して多系統群とされる。
これらの化石鋏角類とカブトガニ類の主流な系統仮説の変化は、大まかに次の通りに挙げられる。
- ウミサソリ類(広翼類、ウミサソリ目/広翼目 Eurypterida)
- ウミサソリ類とカブトガニ類はかつて近縁と考えられ、共に節口類を構成する2つの大群として知られていた。しかしウミサソリ類は、主に精莢の受け渡しに適した生殖器官を基に、20世紀後期以降ではカブトガニ類よりも、クモガタ類との類縁関係(共に Sclerophorata をなす)の方が有力視されるようになっている。
- カスマタスピス類(カスマタスピス目 Chasmataspidida)
- カスマタスピス類は種類によりカブトガニ類とウミサソリ類のいずれかによく似ている。かつて独特なカブトガニ類の一群とも考えられていたが、1990年代後期以降ではカブトガニ類でない独立群として区別されるようになった。カスマタスピス類を多系統群とし、そのうちカブトガニ類に似たカスマタスピス科はカブトガニ類に、ウミサソリ類に似たディプロアスピス科はウミサソリ類に近縁という説もあったが、確実でなく、特に2010年代以降では、本群は単系統群で、ウミサソリ類やクモガタ類に近縁(共に Dekatriata をなす)という説の方が主流になりつつある。
- ハラフシカブトガニ類(共剣尾類、ハラフシカブトガニ亜目/共剣尾亜目 Synziphosurina)
- ハラフシカブトガニ類は伝統的にカブトガニ類に含まれ、派生的なカブトガニ類(カブトガニ亜目 Xiphosurida)の姉妹群もしくはそれに至る側系統群と考えられた。しかし Lamsdell (2013) が様々なハラフシカブトガニ類の形質を検証し、それを基に他の鋏角類との系統関係を再構成したところを初めとして、ハラフシカブトガニ類はカブトガニ亜目と単系統群になっておらず、むしろ真鋏角類全般と Dekatriata 類(カスマタスピス類・ウミサソリ類・クモガタ類からなる単系統群)のそれぞれの初期系統(ステムグループ)から分岐した雑多な分類群だと示唆される(注釈参照)。これを踏まえて、従来および広義のカブトガニ類は、Prosomapoda(オファコルスとダイバステリウム以外の真鋏角類)から Dekatriata 類を除いた側系統群であり、狭義のカブトガニ類は、ハラフシカブトガニ類を除き、カブトガニ亜目とそれに類縁するいくつかの属(ルナタスピス、Kasibelinurus など)のみを含も単系統群と再定義された。
下位分類
World Spider Catalog に掲載される化石鋏角類一覧表「A summary list of fossil spiders and their relatives」(最終更新日:2020年1月15日)によると、現存4種と化石100種以上の広義のカブトガニ類が記載されており、化石種の中で80種以上は狭義のカブトガニ類に属する。
広義のカブトガニ類よりも基盤的な真鋏角類オファコルスとダイバステリウム、および Dekatriata に含まれるとされる Winneshiekia と Houia は、便宜上にハラフシカブトガニ類として広義のカブトガニ類に含まれる場合もある。分類学的位置が議論的で再検証が必要とされる Borchgrevinkium は、Lamsdell (2013) によって暫定的に広義のカブトガニ類としてまとめられる。
狭義のカブトガニ類はほぼ全ての種類がカブトガニ亜目(Xiphosurida)に分類され、そこから2下目2上科5科に細分される。他にもカブトガニ亜目以外の属を含んだKasibelinuridae科はあるが、単系統群をなしていないため再定義が必要と指摘され、それを敢えて採用しない文献もある。
現生種はカブトガニ科に分類され、形態的差異から2亜科3属4種に区別される。人工交雑による実験では、アメリカカブトガニ Limulus polyphemus(アメリカカブトガニ亜科 Limulinae)とアジア産カブトガニ類(カブトガニ亜科 Tachypleininae)では受精が成立せず、アジア産3種の間では交雑卵から孵化する例が見られた。アジア産の3種はマルオカブトガニ Carcinoscorpius rotundicauda(マルオカブトガニ属 Carcinoscorpius)とそれ以外の2種(カブトガニ属)に分けられるが、カブトガニ Tachypleus tridentatus に対してマルオカブトガニよりもミナミカブトガニ Tachypleus gigas の方が生殖的隔離が進んでいることから、マルオカブトガニをカブトガニ属に含める説もある。一方で2012年に発表されたカブトガニ科の分子系統解析では、カブトガニとミナミカブトガニで単系統群を形成し、マルオカブトガニがその姉妹群となるという結果が得られている。
次のリストの中で特記されない限り、分類体系・シノニム・地質時代とも、狭義のカブトガニ類 Xiphosura sensu stricto は Lamsdell (2020) に、その他の広義のカブトガニ類は Dunlop et al. (2020) に基づく。属より上位の分類群は太字、絶滅群は「†」、ジュニアシノニムは「=」、伝統的にハラフシカブトガニ類 Synziphosurina に分類されたものは「*」で示す。
- Xiphosura sensu lato(オルドビス紀 - 現世):広義のカブトガニ類。Dekatriataを除いた側系統群。
- Prosomapoda(Dekatriataを除く)
- (属)Anderella †(石炭紀)*
- (属)Borchgrevinkium †(デボン紀)
- (属)Camanchia †(シルル紀)*
- (属)Legrandella †(デボン紀)*
- (属)Maldybulakia (=Lophodesmus)†(デボン紀)
- ヴェヌストゥルス属 Venustulus †(シルル紀)*
- (属)Willwerathia †(デボン紀)*
- ウェインベルギナ科 Weinberginidae †(デボン紀)*
- ウェインベルギナ属 Weinbergina †(デボン紀)*
- Planaterga(Dekatriataを除く)
- (属)Bembicosoma †(シルル紀)*
- (属)Bunaia †(シルル紀)*
- (科)Pseudoniscidae †(シルル紀)*
- (属)Cyamocephalus †(シルル紀)*
- (属)Pseudoniscus (=Neolimulus)†(シルル紀)*
- (科)Bunodidae †(シルル紀)*
- (属)Bunodes (=Exapinurus)†(シルル紀)*
- リムロイデス属 Limuloides (=Hemiaspis)†(シルル紀)*
- (属)Pasternakevia †(シルル紀)*
- Xiphosura sensu stricto (オルドビス紀 - 現世):狭義のカブトガニ類。
- (属)Pickettia †(デボン紀)
- (属)Patesia †(デボン紀)
- (科)Kasibelinuridae †(オルドビス紀 - デボン紀):カブトガニ亜目 Xiphosurida を除いた側系統群。
- (属)Kasibelinurus †(デボン紀)
- ルナタスピス属 Lunataspis †(オルドビス紀):既知最古の狭義のカブトガニ類。
- カブトガニ亜目 Xiphosurida(石炭紀 - 現世)
- (上科・科未定)
- (属)Albalimulus †(石炭紀)
- (属)Sloveniolimulus †(三畳紀)
- (属)Xiphosuroides †(石炭紀)
- ヒメカブトガニ下目 Bellinurina †(石炭紀 - ペルム紀)
- (科)Bellinuridae (=Euproopidae, Liomesaspididae)†(石炭紀 - ペルム紀)
- (属)Anacontium †(ペルム紀)
- (属)Andersoniella †(石炭紀)
- (属)Alanops †(石炭紀)
- (属)Anacontium †(ペルム紀)
- (属)Belinurus (=Bellinurus, Steropsis, Koenigiella, Macrobelinurus, Parabelinurus)†(石炭紀)
- ユウプループス属 Euproops (=Prestwichia, Prestwichianella)†(石炭紀)
- (属)Koenigiella †(石炭紀)
- (属)Liomesaspis (=Palatinaspis)†(石炭紀 - ペルム紀)
- (属)Patesia †(石炭紀)
- (属)Prestwichianella †(石炭紀)
- (属)Pringlia †(石炭紀)
- (属)Prolimulus †(石炭紀)
- (属)Stilpnocephalus †(石炭紀)
- (科)Bellinuridae (=Euproopidae, Liomesaspididae)†(石炭紀 - ペルム紀)
- カブトガニ下目 Limulina(石炭紀 - 現世)
- (属)Bellinuroopsis (=Neobelinuropsis)†(デボン紀)
- (科)Rolfeiidae †(石炭紀)
- (属)Rolfeia †(石炭紀)
- (上科)Paleolimuloidea †(石炭紀 - ジュラ紀)
- コダイカブトガニ科 Paleolimulidae (=Moravuridae)†(石炭紀 - ジュラ紀)
- (属)Moravurus †(石炭紀)
- (属)Norilimulus †(石炭紀)
- パレオリムルス属 Paleolimulus †(石炭紀 - ペルム紀)
- (属)Xaniopyramis †(石炭紀 - ペルム紀)
- コダイカブトガニ科 Paleolimulidae (=Moravuridae)†(石炭紀 - ジュラ紀)
- カブトガニ上科 Limuloidea(石炭紀 - 現世)
- (属)Valloisella †(石炭紀)
- オーストロリムルス科 Austrolimulidae (=Dubbolimulidae)†(石炭紀 - 三畳紀)
- オーストロリムルス属 Austrolimulus †(三畳紀)
- (属)Batracholimulus †(三畳紀)
- (属)Boeotiaspis †(石炭紀)
- (属)Dubbolimulus†(三畳紀)
- (属)Limulitella (=Limulites)†(三畳紀 - ジュラ紀)
- (属)Panduralimulus †(ペルム紀)
- (属)Psammolimulus †(三畳紀)
- (属)Shpineviolimulus †(ペルム紀)
- (属)Tasmaniolimulus †(三畳紀)
- (属)Vaderlimulus †(三畳紀)
- カブトガニ科 Limulidae(三畳紀 - 現世):現存する唯一の科。
- (属)Allolimulus †(ジュラ紀)
- (属)Casterolimulus †(白亜紀)
- (属)Crenatolimulus †(ジュラ紀 - 白亜紀)
- (属)Heterolimulus †(三畳紀)
- メソリムルス属 Mesolimulus †(三畳紀 - 白亜紀)
- (属)Keuperlimulus †(三畳紀)
- (属)Psammolimulus †(三畳紀)
- (属)Tarracolimulus †(三畳紀)
- (属)Victalimulus †(白亜紀)
- (属)Volanalimulus †(三畳紀)
- (属)Yunnanolimulus †(三畳紀)
- アメリカカブトガニ亜科 Limulinae
- アメリカカブトガニ属 Limulus(白亜紀 - 現世)
- アメリカカブトガニ Limulus polyphemus(現世)
- アメリカカブトガニ属 Limulus(白亜紀 - 現世)
- カブトガニ亜科 Tachypleininae(三畳紀 - 現世):生殖口蓋の内肢は退化的。
- カブトガニ属 Tachypleus(三畳紀 - 現世)
- カブトガニ Tachypleus tridentatus(現世)
- ミナミカブトガニ Tachypleus gigas(現世)
- マルオカブトガニ属 Carcinoscorpius(現世)
- マルオカブトガニ Carcinoscorpius rotundicauda(現世)
- カブトガニ属 Tachypleus(三畳紀 - 現世)
- (上科・科未定)
- Prosomapoda(Dekatriataを除く)
カブトガニ類と誤認された化石
不完全な化石によって知られ、かつてカブトガニ類もしくはその近縁と誤認された古生物はいくつか挙げられる。光楯類(Aglaspidida)はその代表的な一例で、20世紀後期まで、この類は長い間に付属肢が推測的に節口類らしく復元され、カブトガニ類もしくはそれに近縁の鋏角類と誤解釈された。後に付属肢が再検証され、鋏角の代わりに触角があるなど鋏角類らしからぬ特徴をもつと分かり、カブトガニ類の近縁どころか、鋏角類ですらなく、むしろ三葉虫を含んだ別系統(Artiopoda類)のものだと判明した(詳細は光楯類#分類を参照)。光楯類の他、昆虫(Hypatocephala、Permolimulinellaなど)・三葉虫(Hemiaspis)・棘皮動物(Lemoneites)にもカブトガニ類と誤解釈された化石が知られている。後体の局部のみが知られる Kiaeria は長い間にカブトガニ類と思われていたが、Lamsdell (2019) の再検討にむしろカスマタスピス類ではないかと指摘される。断片的な化石のみ知られ、古くからカブトガニ類として記載された Elleria、Archeolimulus と Drabovaspis は Lamsdell (2020) の再検討にカブトガニ類として認められず、特に前者は三葉虫の尾部由来であった可能性が高い。
脚注
注釈
出典
関連項目
- 鋏角類
- 節口綱(腿口綱)
- ハラフシカブトガニ類
- ウミサソリ、カスマタスピス類 - かつてカブトガニ類と近縁と考えられていた節口類。
- 節口綱(腿口綱)
- カブトエビ - しばしばカブトガニ類と混同される。
- 生きている化石